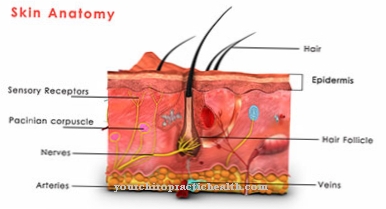
Det Receptorpotentiale er sensoriske cellers respons på en stimulus og svarer normalt til en depolarisering. Det vil det også Generatorpotentiale kaldes og er en direkte konsekvens af de transduktionsprocesser, som receptoren konverterer en stimulus til excitation. Denne proces forstyrres i receptor-associerede sygdomme.
Hvad er receptorpotentialet?

De sensoriske celler i den menneskelige krop kaldes receptorer. Dette er proteiner eller et proteinkompleks, som signalmolekyler binder til. Dette udløser signalprocesser inde i cellerne. Receptorer henter signaler udefra og behandler dem til bioelektrisk excitation. De oversætter stimuli fra miljøet til sprog i centralnervesystemet. Receptorerne er meget specialiserede og er blandt de vigtigste tilfælde af menneskelig opfattelse.
I en upåvirket tilstand har receptorerne et hvilepotentiale. Dette er en spændingsforskel baseret på en ujævn fordeling af natrium- og kaliumioner, der adskiller det intra- og ekstracellulære rum. En indkommende stimulus fra miljøet binder til receptorproteinerne og giver receptoren mulighed for at overskride dens hvilepotentiale. Denne proces kaldes depolarisering. Receptorpotentialet er sensorisk cellers membranelektriske respons på en bestemt stimulus. Nogle forfattere skelner mellem receptorpotentialet og generatorpotentialet. De forstår depolarisationen af en sensorisk neuron som et generatorpotentiale. For dem er et receptorpotentiale imidlertid et potentiale i membranen i receptorcellen.
Funktion & opgave
Receptorpotentialet opstår som et resultat af transduktionsprocessen. Denne proces svarer til konvertering af stimulusenergier til kroppens egen og derfor bearbejdelige ophidselse.
I forbindelse med denne konvertering spiller begrebet signalkaskade en stor rolle. I en vis grad følger de individuelle sensoriske celler forskellige veje til stimulusbehandling og -transduktion. Trinene til binding, transformation, transmission og regenerering er fælles for dem. Depolarisering af sensorisk celle er også et almindeligt trin. Øjets fotoreceptorer er en undtagelse. Lys som en tilstrækkelig stimulus forårsager hyperpolarisering i dem.
Det normale tilfælde er imidlertid depolarisering. Det finder sted i forhold til den respektive styrke af den modtagne stimulus. Afhængig af stimulansstyrken åbnes de membranbaserede kationkanaler som et resultat af ændringer i grundspændingen mellem det intra- og ekstracellulære rum. På denne måde genereres et stimulusgrænseafhængigt handlingspotentiale i receptorens affinitet.
Afference er nervevævet, der er specialiseret i informationsstrømmen. Afferenterne er nervesystemer, der føder ophidselser til det centrale nervesystem.
Forløbet af receptorpotentialet adskiller sig med de respektive receptorer. Potentialet er typisk sammensat af en proportional og en differentiel komponent, så receptorernes stimuleringsrespons er proportional.
Receptorpotentialet skyldes normalt åbningen af de membranbundne natriumkanaler. De frigiver natriumioner i cellen, hvilket forstås som den faktiske excitation. Hyperpolarisationen af fotoreceptorerne sker derimod, når kanalerne er lukket.
Receptorpotentialet er ikke underlagt en alt-eller-intet-lov, men stiger gradvist med stimulansens styrke. Når en bestemt tærskelværdi nås, og tærskelpotentialet således overskrides, genererer den sensoriske celle et handlingspotentiale. Som næsten alle handlingspotentialer følger følelsen af sensoriske celler også en alt-eller-intet-lov og har normalt ingen regenerativ ildfast periode.
Sygdomme og lidelser
Gruppen af receptorassocierede sygdomme påvirker excitationsprocesserne i receptorceller. Dette har også indflydelse på receptorpotentialet. I de senere år har medicinsk forskning opdaget forskellige receptormutationer. Disse mutationer er nu forbundet med en lang række arvelige og somatiske sygdomme.
I receptorassocierede sygdomme er receptorerne defekte. Af denne grund er de ikke længere i stand til at binde sig til signalmolekyler, behandle signaler tilstrækkeligt eller videresende signaler. I tilfælde af andre sygdomme fra denne gruppe kan signaltransduktion næppe eller slet ikke. Andre mutationer kan generelt lade visse receptorer mangle eller inkorporere dem forkert i membranen.
De fleste receptorassocierede sygdomme er ikke forårsaget af receptorerne selv, men af autoantistoffer. Disse autoimmune sygdomme angriber sensoriske celler med deres autoantistoffer og forårsager betændelse. I løbet af denne betændelse ødelægges receptorens indre strukturer, og sensoriske celler mister deres funktionalitet.
Eksempler fra denne gruppe af sygdomme er myasthenia gravis og Lambert-Eaton syndrom. Myasthenia gravis er en autoimmun muskelnervesygdom. Lambert-Eaton-syndrom ligner dette fænomen, men er langt mere almindeligt end myasthenia gravis.
Sygdomme med receptordefekter differentieres efter deres strukturelle klasse. I tilfælde af ionkanalsygdomme forstyrres for eksempel ionekanalernes neuronstruktur og dermed den biokemiske excitabilitet af receptorerne.
Foruden gruppen af receptorassocierede sygdomme kan psykotropiske medikamenter også have en indflydelse på receptorernes signalkaskade. I dette tilfælde virker deres aktive ingredienser direkte på receptorer og efterligner funktionen af den respektive neurotransmitter for at være i stand til at binde til den tilsvarende receptor. Andre psykotropiske lægemidler blokerer receptorerne for fysiologiske neurotransmittere. De beskrevne virkninger af forskellige psykotropiske lægemidler anvendes i moderne medicin specifikt til at påvirke receptoraktiviteter.












.jpg)